100%
4
Rated
Attempts
127
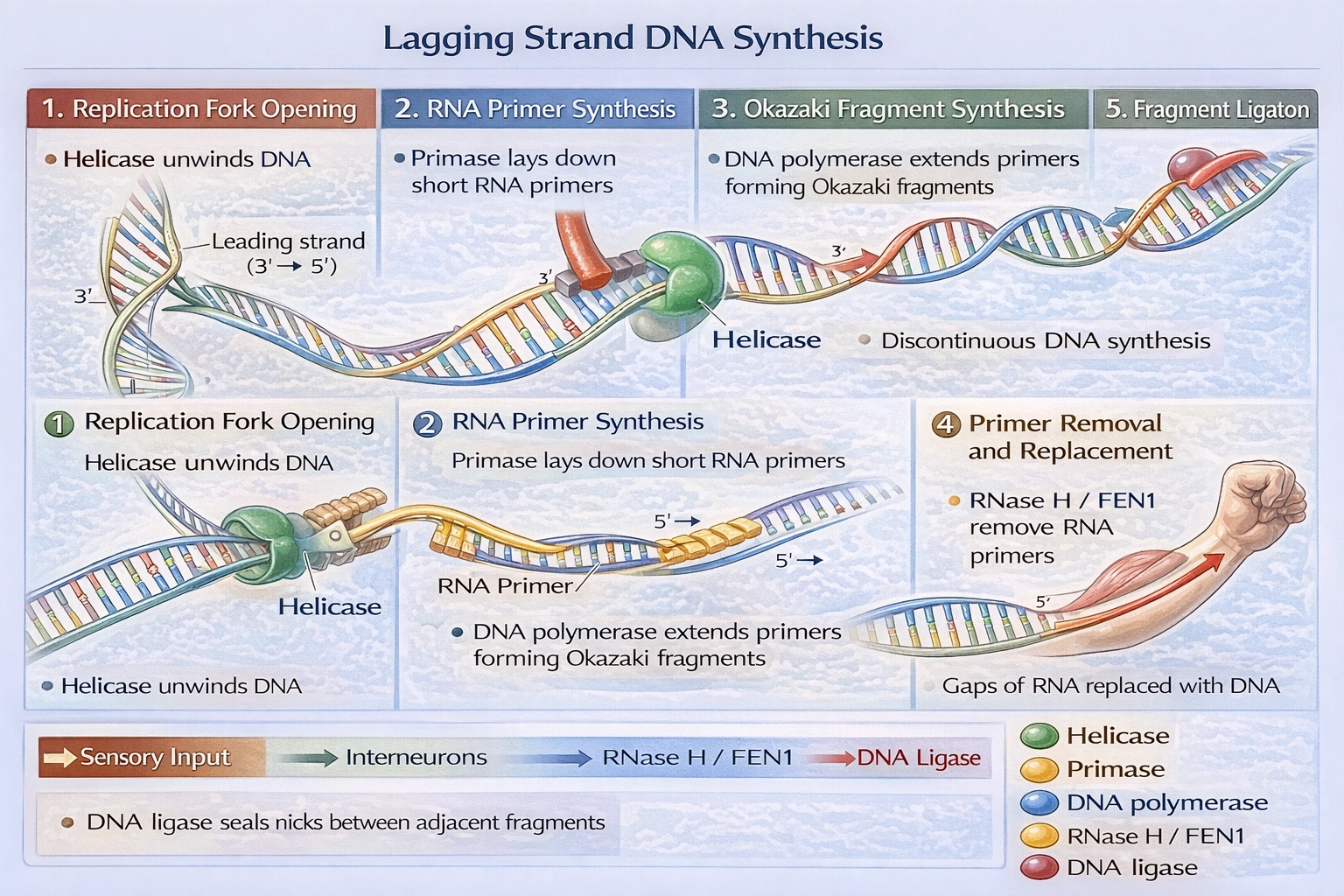
Describe the synthesis of the lagging strand of DNA.
-
A
DNA polymerases synthesize DNA continuously after initially attaching to a primase.
-
B
DNA polymerases synthesize DNA discontinuously in pieces called Okazaki fragments after initially attaching to primases.
-
C
DNA polymerases synthesize DNA discontinuously in pieces called Okazaki fragments after initially attaching to RNA primers.
-
D
DNA polymerases synthesize DNA discontinuously in pieces called Okazaki fragments which are joined together in the end by a DNA helicase.
A
DNA polymerases synthesize DNA continuously after initially attaching to a primase.
B
DNA polymerases synthesize DNA discontinuously in pieces called Okazaki fragments after initially attaching to primases.
C
DNA polymerases synthesize DNA discontinuously in pieces called Okazaki fragments after initially attaching to RNA primers.
D
DNA polymerases synthesize DNA discontinuously in pieces called Okazaki fragments which are joined together in the end by a DNA helicase.
Related Questions
Top Picks
Using anatomical terms, what is the relationship of the sternum relative to the deltoid?
Ligaments connect what?
Identify the correct sequence of the 3 primary body planes as numbered 1, 2, and 3 in the above image.
Which of the following is NOT a major function of the respiratory system in humans?
What is the major difference between somatic and germline mutations?What is the major difference between somatic and germline mutations?
Available FREE Test
Sets
Available Test
Sets